
Dual environmental thresholds and indications of reversible growth-mode regulation in Adenium dhofarense under Dhofar’s hyper-seasonal climate
Author: TAKUYA KAGAWA
Affiliation: Independent Researcher (Succulent Saika, Japan)
ORCID:https://orcid.org/0009-0008-5006-3292
DOI:https://doi.org/10.51094/jxiv.2234
Abstract
Adenium dhofarense is endemic to the Dhofar region of southern Oman, where it experiences an extreme hyper-seasonal climate: a prolonged cool, clouded monsoon (khareef) followed by a brief post-monsoon interval of intense heat and high irradiance. We monitored 20 seedlings over 216 days under outdoor conditions in Japan, documenting caudex growth across varying temperature and light regimes. Under conditions adequate for congeners, A. dhofarense exhibited severe growth suppression. Rapid growth commenced only when high temperature (≥35°C) and high daily light integral (≥23 mol m⁻² day⁻¹) co-occurred, producing 3.5-fold growth acceleration. These thresholds closely match Dhofar’s post-monsoon environmental conditions (40–45°C, estimated 25–31 mol m⁻² day⁻¹). The abrupt, reversible transition between slow-growth and rapid-growth states—triggered strictly by dual-threshold attainment—suggests a potential shift between distinct metabolic modes, possibly involving facultative engagement of C₃-like photosynthesis from a CAM-like default state. This interpretation, while requiring direct physiological validation, is consistent with adaptation to Dhofar’s narrow annual growth window. The strict environmental dependency renders A. dhofarense vulnerable to climate-driven alterations in monsoon timing or intensity, with direct implications for conservation of this endemic species in one of Arabia’s most climatically unique ecosystems.
Introduction
Adenium species are widely cultivated as drought-adapted ornamentals whose early growth dynamics remain insufficiently quantified under controlled horticultural conditions. Across commonly reported cultivation settings—characterized by moderate temperatures (25–35 °C) and adequate light availability consistent with the high-irradiance environments these taxa naturally occupy—seedlings of A. obesum, A. somalense, A. arabicum, and related congeners are typically observed to exhibit progressive caudex thickening during early development. These qualitative observations, while variable among growers and environmental regimes, provide a practical baseline for interpreting species-level differences in biomass allocation and early caudex development. However, the extent to which these observations reflect intrinsic physiological constraints rather than environmental artifacts remains unresolved, particularly for species originating from hyper-arid regions where maximal solar irradiance is the ecological norm.
Adenium dhofarense Rzepecky (2015) is restricted to the coastal escarpment of Dhofar, southern Oman, where it is exposed to a climatic regime unique within the genus. During the khareef monsoon (June–September), persistent orographic cloud cover produces cool conditions (22–28 °C) and severely reduced direct solar irradiance, resulting in a prolonged period of limited photosynthetic opportunity. After the monsoon, the environment shifts abruptly to intense heat (40–45 °C maxima) and cloudless skies, creating a narrow post-monsoon interval characterized by maximal light and temperature availability. This sharply pulsed resource regime contrasts with the more predictable rainfall-driven seasonality typical of other Adenium habitats.
Although the species has attracted horticultural interest since its description, A. dhofarense remains physiologically undocumented. Preliminary anecdotal accounts describe unusually slow seedling growth compared with congeners, yet no quantitative analyses exist regarding its environmental thresholds or photosynthetic behavior. This lack of empirical characterization obscures whether the species possesses intrinsic physiological constraints consistent with its highly seasonal native environment.
For comparative context, we also cultivated Adenium somalense Balf.f. (1888) (regarded by some authorities as a synonym of A. obesum, e.g., POWO, Royal Botanic Gardens Kew, but treated here as a distinct entity based on its consistent morphological differentiation and contrasting growth habit), a taxon characterized by narrow, elongate leaves and slender growth habit.
During long-term routine cultivation of multiple Adenium species (over ten years of continuous annual seedling production), A. dhofarense was not initially treated as an experimental subject but simply incorporated into standard sowing batches. Because no physiological or horticultural information was available for the species, seedlings were started under a conservative partial-shade acclimation regime that reliably supports early development in other Adenium taxa. Until late June, no systematic measurements were taken, as no deviation from expected genus-level behavior was anticipated.
This assumption changed abruptly in late June 2025, when several A. dhofarense seedlings exhibited marked stress—most notably leaf abscission and near-complete growth stagnation—despite environmental conditions (moderate temperatures and irradiance) that provide conditions permitting early growth of A. obesum, A. somalense, and A. arabicum. These unexpected responses, unprecedented in more than a decade of comparative seedling experience, motivated a transition from routine cultivation to structured monitoring beginning on 30 June. The study therefore originates from the emergence of anomalous physiological behavior incompatible with the response patterns typically assumed for Adenium.
Here, we document the environmental conditions associated with seedling growth responses in A. dhofarense over 216 days of outdoor cultivation under varying light and temperature regimes, accompanied by a concurrently grown cohort of A. somalense as a congeneric reference. Rather than testing a predefined hypothesis, we assess whether the observed growth patterns in A. dhofarense are consistent with the interpretation that the species exhibits constrained photosynthetic flexibility, requiring the concurrent attainment of high-temperature and high-irradiance thresholds to shift from slow, stress-tolerant growth to rapid biomass accumulation.
We interpret these patterns in the context of Dhofar’s sharply seasonal climate and discuss potential implications for metabolic strategy within Apocynaceae.
Materials & Methods
Study system and cultivation origin
Seeds of A. dhofarense were germinated on 16 April 2025 in 6‑cm plastic pots containing pH‑adjusted peat moss as the sole substrate, under outdoor conditions in Amagasaki, Japan (34.7°N, 135.4°E). Initial placement under partial shade (~50% natural shading by adjacent structures) followed a conservative acclimatization protocol routinely employed for rare succulent taxa in our facility, in which early-stage seedlings are gradually transitioned to full sun once vigorous growth is established. This protocol has reliably supported early development in A. obesum, A. somalense, A. arabicum, and related taxa over more than a decade of continuous cultivation.
Seeds of A. somalense were germinated on 24 June 2025 under identical substrate and container conditions, providing a congeneric reference cohort 69 days younger than A. dhofarense. Both species were subjected to identical outdoor conditions (natural rainfall, full sun exposure) from 1 July onward.
Transition from routine to structured monitoring
Through late June, no systematic quantitative measurements were conducted because A. dhofarense had been incorporated into routine seedling production rather than treated as an experimental subject. During this period, the cohort exhibited generally slow early development and suboptimal vigor relative to long-term experience with congeners. A subset of individuals showed leaf abscission and apparent growth arrest under environmental conditions (maximum ~33 °C with incident light levels generally permitting viable growth in other Adenium species) that normally support stable growth in other Adenium species. Recognition of these atypical responses prompted a management change: all seedlings were moved to full-sun exposure on 1 July, and continuous qualitative observation was initiated.
By late July, the cohort displayed consistent recovery trends under full sun. On 30 July (Day 105), a single representative plant (designated D‑1) was selected for detailed quantitative documentation. D‑1 showed a recovery trajectory typical of the healthier portion of the cohort and had not undergone complete defoliation during the June stress period.
Quantitative measurements were not collected at fixed intervals. Instead, the plants were observed daily, and measurements of D‑1 (caudex diameter, leaf dimensions) were recorded opportunistically when discernible morphological change occurred. This event‑triggered recording approach resulted in irregular measurement spacing from Day 105 through Day 216 (18 November 2025), reflecting operational practicality and the growth characteristics of the species. Cohort‑level observations were concurrently maintained to contextualize the responses of the focal individual.
Environmental data acquisition
Air temperature and solar irradiance data were obtained from the NEDO METPV database (Kobe station, 34.70°N, 135.21°E), located approximately 20 km from the cultivation site. Horizontal global irradiance (W m⁻²) was converted to estimated photosynthetic photon flux density (PPFD, µmol m⁻² s⁻¹) by assuming that ~45% of total irradiance falls within the PAR range and applying a standard quantum conversion factor (1 W m⁻² PAR ≈ 4.6 µmol m⁻² s⁻¹), yielding PPFD ≈ irradiance × 2.1.
Daily light integral (DLI, mol m⁻² day⁻¹) was approximated from hourly irradiance data using a simplified expression that accounts for the typical bell-shaped distribution of daylight irradiance:
$$
\mathrm{DLI} \approx \frac{\mathrm{PPFD}_{\text{est}} \times \mathrm{sunshine\_hours} \times 0.5}{1000}
$$
The coefficient 0.5 reflects the mean‑to‑peak ratio of daytime irradiance and is commonly applied when only daily irradiance summaries are available. These values provide relative indices of daily light availability but are not intended as precise integrals; they do not capture microclimatic variation specific to the cultivation site (e.g., localized shading or thermal buffering).
Measurements
Caudex diameter was measured with a digital caliper (±0.01 mm precision) at the widest point of the stem base. Leaf dimensions (length and width of the largest fully expanded leaf) were obtained using the same instrument. Photographs were taken under natural light from standardized angles whenever notable phenological changes occurred, including leaf abscission, recovery, and the onset of seasonal dormancy.
Quantitative measurements were not performed at fixed intervals. Instead, observations were made continuously as part of routine cultivation, and measurements were recorded when clear morphological change was detected or when growth trajectories required documentation. After 30 July, one representative individual of A. dhofarense (D‑1) was selected for consistent tracking because its morphology and recovery pattern were typical of the cohort.
A. somalense, which was being cultivated concurrently under identical outdoor conditions, was incorporated as a comparative reference species. Measurements for the representative A. somalense individual were collected on the same days as those of D‑1 to allow direct comparison of developmental responses despite the difference in germination timing.
Results
Early seedling responses under partial shade and moderate temperatures
During the initial 76–98 days after germination (16 April–30 June), A. dhofarense seedlings grew under partial shade (~50% reduction in incident light) with maximum daily temperatures of 30–33 °C. Across this period, seedlings exhibited minimal caudex expansion and poor leaf retention relative to long‑term baseline expectations for congeners. Several individuals displayed complete or near‑complete leaf abscission, and no seedling demonstrated the steady low‑to‑moderate growth typically observed in A. obesum, A. somalense, or A. arabicum under equivalent environmental conditions.
Despite adequate hydration and absence of visible pathogen damage, growth remained stagnant, indicating that the environmental regime—normally sufficient for early Adenium seedling development—was inadequate for A. dhofarense.
In contrast, the A. somalense cohort (germinated 24 June) showed no comparable suppression of early growth under the same partial‑shade transitional conditions, consistent with the broader thermal and irradiance tolerance documented for this species.
Recovery under full sun: irradiance alone is insufficient
Following recognition of atypical stress responses, all seedlings were moved to full sun on 1 July (Day 76). Over the subsequent 3–4 weeks, qualitative improvements in vigor were observed across the A. dhofarense cohort: leaf retention increased, new leaf emergence became more frequent, and overall plant turgor improved. However, caudex enlargement remained limited.
The focal plant D‑1 showed only modest diameter increases through late July despite continuous exposure to midday PPFD levels exceeding those previously experienced under partial shade. On 30 July (Day 105), caudex diameter was 9 mm, representing minimal growth since the transfer to full sun.
These observations indicate that increased irradiance alone did not induce substantial growth recovery. In the absence of high temperatures, irradiance up to the typical midsummer range in Amagasaki (DLI ~17–19 mol m⁻² day⁻¹ during this period) was insufficient to shift the species into a rapid growth mode.
By contrast, A. somalense maintained stable growth under the same conditions, reinforcing that A. dhofarense possesses a narrower effective environmental envelope.
Rapid growth occurs only when high temperature and high irradiance coincide
A marked acceleration in growth was first detected between late July and mid‑September, corresponding to a period during which maximum daily temperatures consistently exceeded 35 °C and estimated DLI frequently surpassed 23 mol m⁻² day⁻¹.
Under these combined conditions, D‑1 exhibited sustained increases in caudex diameter, expanding from 9 mm (30 July, Day 105) to 23.8 mm (18 September, Day 156)—a 164% increase over 51 days (mean growth rate ~0.29 mm day⁻¹). This rate was approximately 3.5‑fold higher than the pre‑acceleration phase (Days 76–105: ~0.08 mm day⁻¹).
The shift in growth behavior was abrupt and temporally aligned with the joint occurrence of elevated temperature and irradiance; neither variable alone appeared sufficient. For example:
- Days with high irradiance but temperatures ≤33 °C did not induce rapid
growth. - Days with temperatures ≥35 °C but reduced irradiance (<20 mol m⁻² day⁻¹
DLI) were also not associated with accelerated diameter increase.
This pattern indicates that A. dhofarense requires a dual‑threshold environmental window to transition from a slow, stress‑tolerant mode to a rapid growth mode, consistent with the hypothesis of constrained photosynthetic flexibility. The concurrent A. somalense individual, despite being 69 days younger, maintained continuous growth across the full range of July–September conditions, further highlighting the unusually strict environmental dependence of A. dhofarense.
Seasonal decline and individual variation in dormancy response
As seasonal temperatures declined below 30–32 °C in mid‑October and DLI values decreased with shortening day length, D‑1 exhibited a marked reduction in growth rate, transitioning toward dormancy. By 18 November (Day 216), when maximum temperatures had fallen to 21 °C and DLI to ~16 mol m⁻² day⁻¹, D‑1 had undergone complete leaf abscission, indicating entry into a dormant state. The cessation of rapid growth aligned precisely with the loss of environmental conditions that had previously supported accelerated caudex expansion.
Neither moderate irradiance nor residual late‑autumn warmth was sufficient to maintain high growth rates once maximum temperatures no longer exceeded 35 °C and DLI fell below 23 mol m⁻² day⁻¹.
Notably, dormancy onset varied substantially among individuals within the cohort. While D‑1 and several other plants had fully defoliated by mid‑November, a subset of individuals retained green, turgid leaves and showed no signs of dormancy entry despite identical environmental conditions. This intraspecific variation suggests that dormancy timing in A. dhofarense is not strictly governed by external cues alone, but may reflect genetic diversity or developmental state differences among seedlings.
Visual documentation of this dormancy variation is presented in Figure 3. By mid‑November, when dual‑threshold conditions had been absent for over one month, approximately half of the cohort—including D‑1—had undergone complete defoliation and exhibited dormant caudex appearance. The remaining individuals, however, retained fully expanded, photosynthetically active leaves with no visible signs of metabolic down‑regulation. This marked intraspecific heterogeneity occurred despite uniform cultivation conditions, indicating that dormancy timing in A. dhofarense is not solely governed by external environmental cues but is modulated by intrinsic factors that vary among seedlings.
This pattern suggests that A. dhofarense reverts to a low‑energy metabolic mode outside of the narrow window in which high temperature and high irradiance co‑occur. The observed seasonal trajectory is consistent with a strategy in which rapid biomass accumulation is restricted to limited periods of coincident environmental maxima.
In contrast, the younger A. somalense reference continued modest growth into late autumn under the same conditions, with no individuals showing dormancy by mid‑November, indicating that the strict environmental dependence and early dormancy onset observed in A. dhofarense are not generalized features of the genus but rather species‑specific physiological constraints.
Discussion
Summary of key findings and environmental threshold model
In this study, we demonstrated that Adenium dhofarense seedlings require the concurrent attainment of high temperature (≥35 °C maximum) and high daily irradiance (estimated DLI ≥23 mol m⁻² day⁻¹) to initiate rapid caudex growth. Under partial shade or moderate temperature alone, growth remained minimal, and many individuals experienced leaf abscission. The data reveal a dual‑threshold environmental window governing seedling growth dynamics: outside this window, A. dhofarense appears to revert to a low‑energy, slow‑growth mode.
These results contrast sharply with the behavior of the congeneric A. somalense, which maintained continuous growth across a broader and less extreme range of temperature and light conditions. Thus, A. dhofarense exhibits constrained photosynthetic and growth flexibility relative to congeners, a pattern that cannot be attributed solely to cultivation artifacts but appears to reflect inherent physiological constraints.
Ecological interpretation in light of native habitat seasonality
The strict dual‑threshold requirement aligns closely with the native climatic regime of the Dhofar escarpment. This region is characterized by the khareef monsoon (June–September), during which persistent orographic cloud cover produces cool temperatures (22–28 °C) and severely reduced direct solar irradiance, followed by a brief post‑monsoon interval of intense heat (40–45 °C maxima) and cloudless skies (Horisk et al., 2023, 2024). In its natural habitat, A. dhofarense likely experiences a narrow but intense favorable season, during which rapid biomass accumulation must occur before the onset of the prolonged dry season.
The observed physiological strategy—slow or near‑dormant growth outside of the optimal window, rapid growth only under combined environmental maxima—may represent an adaptive trade‑off, allowing survival through extended unfavorable periods while maximizing carbon gain during the brief favorable interval. This interpretation supports the view that the dual‑threshold growth requirement is not a cultivation artifact but an evolved adaptation to the sharply pulsed resource availability characteristic of its endemic environment (Horisk et al., 2024).
Implications for metabolic strategy: hypothesis of constrained CAM–C₃ flexibility
Although we do not present direct physiological data (e.g., gas‑exchange measurements, diel titratable acidity, or carbon isotope discrimination) to confirm photosynthetic mode, the growth behavior observed in A. dhofarense is consistent with a metabolic strategy in which the plant defaults to a low‑energy, CAM‑like state under suboptimal conditions and transitions to a high‑activity, C₃‑like mode only under optimal external cues. Because no peer‑reviewed studies have conclusively demonstrated the predominant photosynthetic pathway or CAM reversibility in Adenium species, this interpretation does not rely on an assumed “baseline C₃ with facultative CAM” framework for the genus. Instead, our hypothesis is strictly comparative: relative to the typical growth responses reported for congeners under similar conditions, A. dhofarense appears to show an unusually strong dependency on simultaneous high irradiance and high temperature to achieve sustained daytime carbon gain.
This hypothesis is consistent with the observed growth patterns: the extremely slow early growth under moderate conditions (suggestive of limited CO₂ uptake via nocturnal stomatal opening), the abrupt acceleration when dual thresholds are met (consistent with daytime stomatal opening and higher photosynthetic rates; cf. Yamori et al., 2014), and the rapid reversion to dormancy when conditions decline (suggestive of re‑engagement of water‑conserving CAM mode). Such a strategy would represent a novel adaptation within Adenium, potentially convergent with obligate CAM taxa in other succulent lineages.
Intraspecific variation in dormancy onset: ecological implications
The observed variation in dormancy timing among individuals under identical environmental conditions suggests that intrinsic factors—possibly genetic diversity or ontogenetic asynchrony—influence the threshold for metabolic down‑regulation and leaf abscission. While focal individual D‑1 and several cohort members had fully defoliated by mid‑November, a substantial subset retained green, turgid leaves with no apparent dormancy entry. This intraspecific variability may reflect a bet‑hedging strategy, allowing at least some seedlings to exploit variable or extended favorable windows if environmental conditions fluctuate unpredictably—a pattern commonly observed in species endemic to hyper‑seasonal environments (Horisk et al., 2023).
From a conservation perspective, this variability indicates that A. dhofarense populations may harbor considerable physiological diversity, which could be critical for persistence under climate variability. Moreover, the apparent reliance on narrowly defined high‑temperature and high‑irradiance conditions for sustained growth suggests that populations occupying microhabitats with differing thermal or light regimes may experience divergent selective pressures, potentially shaping local physiological differentiation relevant to long‑term species resilience.
Limitations, physiological validation priorities, and broader implications
This study is limited by its reliance on morphological proxies (caudex diameter, leaf phenology) rather than direct measurements of photosynthetic physiology. The central claim of this work—that A. dhofarense exhibits reversible shifts between slow, CAM‑like metabolism and rapid, C₃‑like metabolism in response to environmental conditions—remains a hypothesis requiring physiological validation. The dual‑threshold environmental window (≥35 °C maximum temperature and ≥23 mol m⁻² day⁻¹ DLI) represents the conditions under which this metabolic transition appears to occur, but the precise numerical values should be interpreted as approximate indicators rather than strict physiological constants.
The estimation of DLI from horizontal global irradiance obtained from a meteorological station on the order of 20 km distant introduces uncertainty in absolute values; however, the core evidence lies not in the exact threshold magnitudes but in the demonstrable reversibility of the growth response across distinct environmental regimes. The transition from minimal growth under moderate conditions, to rapid growth when both temperature and irradiance maxima are reached, and back to dormancy when conditions decline, constitutes a reproducible phenological pattern that is inconsistent with constitutive C₃ metabolism but consistent with environmentally triggered metabolic switching.
Gas‑exchange measurements and diel titratable acidity assays planned for 2026 will directly test whether the observed growth transitions correspond to shifts in stomatal behavior and CO₂ uptake timing. Controlled‑environment experiments comparing plants maintained at 25 °C versus 35 °C under standardized irradiance will allow quantification of net photosynthetic rates, stomatal conductance, and nocturnal acid accumulation, following established protocols for CAM assessment (Winter & Holtum, 2014; Yamori et al., 2014). Complementary stable‑carbon isotope analysis (δ¹³C) of leaf tissue produced under contrasting environmental conditions will provide time‑integrated evidence of photosynthetic pathway utilization. Until these data are obtained, the metabolic interpretation presented here should be regarded as a working hypothesis grounded in phenological observation rather than confirmed physiology.
Nonetheless, the phenological evidence for reversible, threshold‑dependent growth mode switching in A. dhofarense is robust and represents a novel pattern within the genus. If confirmed physiologically, this species would exemplify a strategy in which constitutive or near‑constitutive CAM serves as the default state, with facultative engagement of C₃‑like metabolism restricted to brief windows of extreme environmental favorability—a departure from the physiological scenarios discussed by Winter & Holtum (2014), in which CAM expression is typically framed in relation to environmental stress responses, rather than as a default metabolic state. Such a strategy would be consistent with adaptation to the hyper‑seasonal climate of the Dhofar escarpment, where extended periods of marginal conditions (cool, clouded khareef monsoon) are interrupted by short, intense favorable intervals of extreme heat and high irradiance (Horisk et al., 2024). Whether similar threshold‑dependent metabolic flexibility exists in other Adenium species, and whether it represents a shared ancestral trait or independent adaptation to extreme environments, remains to be determined through comparative physiological studies.

Figure 1. Phenological transitions in Adenium dhofarense seedlings across environmental regimes.
(A) Initial stress under partial shade (30 June 2025, Day 75): severe leaf abscission (white arrow indicates nearly defoliated individual). During this phase, expansion of even a single leaf required several weeks, underscoring the extremely slow pace of growth prior to the rapid phase.
(B) Early recovery under full sun but moderate temperature (22 July 2025, Day 97): leaf retention improved but caudex growth remained minimal.
(C) Cohort comparison showing delayed development of A. dhofarense (left) relative to A. somalense (right, germinated 69 days later) under identical conditions.
(D) Focal individual D‑1 at onset of rapid growth phase (30 July 2025, Day 105): caudex diameter 9 mm, selected as the largest individual in the cohort.
(E) D‑1 after one week under dual‑threshold conditions (6 August 2025, Day 112): marked increase in leaf number, area, and thickness, representing growth acceleration exceeding the entire preceding month.
(F) Continued rapid expansion under sustained high temperature and high irradiance (19 August 2025, Day 125): caudex diameter 12 mm, demonstrating the abrupt shift to high‑activity growth mode.
Substrate transition from peat moss (A–B) to high‑drainage mineral mix (D–F) occurred during the recovery phase. All images were taken under identical outdoor cultivation in Amagasaki, Japan.
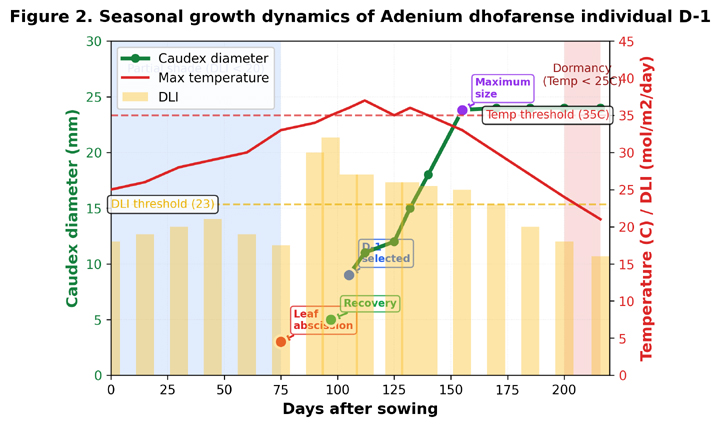
Figure 2. Seasonal growth dynamics of Adenium dhofarense individual D‑1.
Caudex diameter (green line, left axis) plotted against days after sowing, with maximum daily temperature (red line, right axis) and estimated daily light integral (DLI, yellow bars). Key phenological events are annotated: leaf abscission under partial shade (Day ~75), selection of D‑1 for detailed tracking (Day 105), recovery phase (Days 97–105), and rapid growth phase (Days 105–156) coinciding with temperatures ≥35 °C and DLI ≥23 mol m⁻² day⁻¹. Blue shaded region indicates partial‑shade period; unshaded region indicates full‑sun exposure. Pink shaded region indicates dormancy onset (temperature <25 °C). Growth rate increased approximately 3.5‑fold when both temperature and irradiance thresholds were exceeded simultaneously, demonstrating obligate dual‑threshold dependence.

Figure 3. Dormancy response of Adenium dhofarense seedlings under declining late‑autumn conditions.
Photographed 18 November 2025 (Day 216), when maximum daily temperature had fallen to 21 °C and estimated DLI to ~16 mol m⁻² day⁻¹. Under these conditions—well above the temperatures at which other Adenium species typically remain active—approximately half of the A. dhofarense cohort exhibited complete leaf abscission and entry into dormancy (left, focal individual D‑1). In contrast, a subset of individuals retained green, turgid leaves (right) despite identical environmental conditions. This early onset of dormancy at moderate temperatures highlights the unusually strong dependence of A. dhofarense on high temperature and high irradiance for sustained growth, distinguishing it physiologically from other congeners. Both plants were maintained under full sun and natural rainfall throughout the observation period.
References
- Balfour, I. B. (1888). Adenium somalense Balf.f.In Transactions of the Royal Society of Edinburgh (Vol. 31, p. 162).Edinburgh: Royal Society of Edinburgh.
- Horisk, K. E., Ivory, S. J., Lézine, A.-M., Watrin, J., & Renson, V. (2023). Vegetation dynamics in Dhofar, Oman, from the Late Holocene to present inferred from rock hyrax middens.
Quaternary Research, 116, 12–29.
https://doi.org/10.1017/qua.2023.42 - Horisk, K. E., Ivory, S. J., Lézine, A.-M., Watrin, J., & Renson, V. (2024). Late Holocene hydrologic variability and ecosystem structure from rock hyrax middens in Dhofar, Oman.
Frontiers in Earth Science, 12, 1441323.
https://doi.org/10.3389/feart.2024.1441323 - POWO. Adenium somalense Balf.f. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew.
Available at:
https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:76372-1
(Accessed: December 2025). - Rzepecky, A. (2015). Adenium dhofarense sp. nov.: A long-overlooked species from the central-southern part of Oman.Cactus and Succulent Journal, 87(3), 129–135.
https://doi.org/10.2985/015.087.0306 - Winter, K., & Holtum, J. A. M. (2014). Facultative crassulacean acid metabolism (CAM) plants: Powerful tools for unravelling the functional elements of CAM photosynthesis.
Journal of Experimental Botany, 65(13), 3425–3441.
https://doi.org/10.1093/jxb/eru063 - Yamori, W., Hikosaka, K., & Way, D. A. (2014). Temperature response of photosynthesis in C₃, C₄, and CAM plants: Temperature acclimation and adaptation.
Photosynthesis Research, 119, 101–117.
https://doi.org/10.1007/s11120-013-9874-6
Data Availability
Environmental data (temperature, irradiance) were obtained from the publicly accessible
NEDO METPV database (https://www.nedo.go.jp/library/nissharyou.html). Morphological
measurement data and photographs are available from the author upon reasonable request.